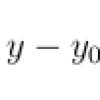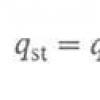Система комплемента: общее представление. Комплемент, состав, основные свойства. Пути активации. Участие комплемента в реакциях иммунитета. РСК, методика ее постановки и практическое использование Что такое комплимент в микробиологии
/ 20
ХудшийЛучший
Комплементом называют сложную систему белков (более 20) сыворотки крови, обладающих ферментативной активностью. Основные 13 компонентов системы комплемента обозначаются буквой С с соответствующим номером (CI, С2, СЗ и т. д.) Они образуются в печени и секретируются макрофагами. Активация системы комплемента протекает классическим и альтернативным путями в виде цепной реакции, управляемой 7-ю регуляторными белками. При этом каждый предыдущий компонент каскада активирует несколько последующих за счет их ферментативного расщепления. Активированные компоненты обозначаются сверху чертой.
Естественный ингибитор комплемента С1 (С 1-ингибитор) тормозит спонтанную активацию Clq компонента.
Классический путь активации Запускается комплексом антиген-антитело в присутствии катионов Са и Mg обычно на поверхности клетки-мишени. Комплекс антиген-антитело связывается с C1q, который присоединяет C1rs, а затем активирует и расщепляет С4 на С4а и С4b. С4b присоединяется либо к С1, либо к поверхности клетки-мишени. Далее к нему присоединяется С2. Он, в свою очередь, расщепляется на С2а и С2b предыдущим компонентом. С2а остается связанным с С4b. Этот комплекс получил название конвертазы классического пути активации комплемента. Она расщепляет СЗ компонент на СЗа и СЗb. СЗb присоединяется к конвертазе классического пути и этот макромолекулярный комплекс активирует компонент С5. Он распадается на С5а и С5b. К С5b на мембране клетки-мишени последовательно присоединяются С6, С7, С8 и С9 компоненты. Комплекс С5b-С9 получил название мембраноатакующего комплекса (МАК). В механизме его литического действия много общего с перфорином. МАК встраивается в мембрану клетки-мишени за счет гидрофобных взаимодействий, образуя трансмембранный канал. Через него в клетку поступают ионы натрия и вода, а выходят ионы калия, что приводит к цитолизу.
Классический путь активации комплемента может запускаться белками А стафилококка, комплексами С-реактивного белка с микробными продуктами и др.
Альтернативный путь активации Комплемента является неспецифическим. Он запускается липополисахаридами клеточной стенки бактерий (эндотоксинами), агрегированными иммуноглобулинами, лекарственными препаратами и т. д. Образующийся при этом СЗb-компонент в присутствии ионов магния связывается с фактором В сыворотки (неактивная сериновая протеаза). На комплекс СЗbВ действует фактор D - активная сывороточная протеаза. Она расщепляет фактор В на Ва и Вb. Образующийся комплекс СЗbВb представляет собой конвертазу альтернативного пути активации. В норме она неустойчива, но стабилизируется белком пропердином (белок Р). Конвертаза альтернативного пути активирует С5 компонент. Дальнейшая активация комплемента не отличается от классического пути. Таким образом, СЗ-компонент является ведущим в активации комплемента по обоим путям, определяя процессы цитолиза. В процессе активации комплемента образуются биологически активные фрагменты. Так, компоненты СЗа и C5a являются анафилатоксинами, действуют на макрофаги, гранулоциты, тучные клетки. Возникающий патологический процесс клинически проявляется аллергическими и псевдоаллергическими реакциями.
При заболеваниях, сопровождающихся образованием иммунных комплексов (аутоиммунные болезни, инфекции), уровень белков комплемента снижается - гипокомплементемия. Уровень комплемента наиболее высок у морских свинок, поэтому их сыворотка крови используется как " комплемент" в серологических реакциях.
Компоненты активированного комплемента связываются с рецепторами комплемента, имеющимися на лейкоцитах: CR1(CD35) - рецептор 1-го типа, связывает СЗb, есть на эритроцитах и лейкоцитах, он же связывает вирус Эпштейна - Барра; CR2(CD21) связывает C3d, имеется на лимфоцитах; CR3(CD11b/CD18) связывает C3bi, экспрессирован на гранулоцитах, участвует в фагоцитозе; CR4 (CD11c/CD18) для СЗа присутствует на фагоцитах.
Взаимодействуя с этими рецепторами клеток продукты активации комплемента стимулируют функции лейкоцитов, запускают воспаление; усиливают противомикробный иммунитет.
Природа и характеристика комплемента . Комплемент является одним из важных факторов гуморального иммунитета, играющим роль в защите организма от антигенов. Комплемент представляет собой сложный комплекс белков сыворотки крови, находящийся обычно в неактивном состоянии и активирующийся при соединении антигена с антителом или при агрегации антигена. В состав комплемента входят 20 взаимодействующих между собой белков, девять из которых являются основными компонентами комплемента; их обозначают цифрами: С1, С2, СЗ, С4... С9. Важную роль играют также факторы В, D и Р (пропердин). Белки комплемента относятся к глобулинам и отличаются между собой по ряду физико-химических свойств. В частности, они существенно различаются по молекулярной массе, а также имеют сложный субъединичный состав: Cl-Clq, Clr, Cls; СЗ-СЗа, СЗЬ; С5-С5а, С5b и т. д. Компоненты комплемента синтезируются в большом количестве (составляют 5-10% от всех белков крови), часть из них образуют фагоциты.
Функции комплемента многообразны: а) участвует в лизисе микробных и других клеток (цитотоксическое действие); б) обладает хемотаксической активностью; в) принимает участие в анафилаксии; г) участвует в фагоцитозе. Следовательно, комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов (например, опухолевых клеток, трансплантата).
Механизм активации комплемента очень сложен и представляет собой каскад ферментативных протеолитических реакций, в результате которого образуется активный цитолитический комплекс, разрушающий стенку бактерии и других клеток. Известны три пути активации комплемента: классический, альтернативный и лектиновый.
По классическому пути комплемент активируется комплексом антиген-антитело. Для этого достаточно участия в связывании антигена одной молекулы IgM или двух молекул IgG. Процесс начинается с присоединения к комплексу АГ+АТ компонента С1, который распадается на субъединицы Clq, Clr и С Is. Далее в реакции участвуют последовательно активированные «ранние» компоненты комплемента в такой последовательности: С4, С2, СЗ. Эта реакция имеет характер усиливающегося каскада, т. е. когда одна молекула предыдущего компонента активирует несколько молекул последующего. «Ранний» компонент комплемента С3 активирует компонент С5, который обладает свойством прикрепляться к мембране клетки. На компоненте С5 путем последовательного присоединения «поздних» компонентов С6, С7, С8, С9 образуется литический или мембраноатакующий комплекс который нарушает целостность мембраны (образует в ней отверстие), и клетка погибает в результате осмотического лизиса.
Альтернативный путь активации комплемента проходит без участия антител. Этот путь характерен для защиты от грамотрицательных микробов. Каскадная цепная реакция при альтернативном пути начинается с взаимодействия антигена (например, полисахарида) с протеинами В, D и пропердином (Р) с последующей активацией компонента СЗ. Далее реакция идет так же, как и при классическом пути - образуется мембраноатакующий комплекс.
Лектиновыи путь активации комплемента также происходит без участия антител. Он инициируется особым маннозосвязывающим белком сыворотки крови, который после взаимодействия с остатками маннозы на поверхности микробных клеток катализирует С4. Дальнейший каскад реакций сходен с классическим путем.
В процессе активации комплемента образуются продукты протеолиза его компонентов - субъединицы СЗа и СЗb, С5а и С5b и другие, которые обладают высокой биологической активностью. Например, СЗа и С5а принимают участие в анафилактических реакциях, являются хемоаттрактантами, СЗb - играет роль в опсонизации объектов фагоцитоза, и т. д. Сложная каскадная реакция комплемента происходит с участием ионов Са 2+ и Mg 2+ .
Московская Государственная Академия Ветеринарной
медицины и Биотехнологии им. К.И.Скрябина
Реферат по иммунологии на тему: «Система комплимента»
Работу выполнила
Котлярова А. Д.
6 группа 3 ФВМ
Работу проверил
Москва 2008 г.
Система комплемента - сложный комплекс белков, представленных главным образом во фракции β-глобулинов, насчитывающий, включая регуляторные, около 20 компонентов, на долю которых приходится 10 % белков сыворотки крови. Комплемент впервые описал Бухнер в 1889 г. под названием «алексин» - термолабильный фактор, вприсутствии которого наблюдается лизис микробов. Свое название (Эрлих, 1895) комплемент получил благодаря тому, что он комплементирует (дополняет) и усиливает действие антител и фагоцитов, защищая организм человека и животных от большинства бактериальных инфекций.
Комплемент представляет собой систему каскадно-действующих пептид-гидролаз, получивших обозначение от С1 до С9. Установлено, что большая часть компонентов комплемента синтезируется гепатоцитами и другими клетками печени (около 90 %, СЗ, С6, С8, фактор В и др.), а также моноцитами/макрофагами (С1, С2, СЗ, С4, С5).
С1-компонент представлен в плазме крови тремя белками(Clq, Clr, С Is).
Наиболее сложной из них является молекула Clq (рис. 1), состоящая из 18 полипептидных цепей трех типов (по 6 цепей А-, В- и С-типов). Все 18 цепей своими коллагеноподобными N-конца-ми (78 остатков аминокислот) образуют канатообразную спирально закрученную структуру, от которой расходятся в разные стороны С-концевые участки цепей (103-108 аминокислотных остатков), завершающиеся глобулярными головками, которые могут взаимодействовать с комплементсвязывающими участками Сн-доменов антител (в составе иммунного комплекса АГ-AT).
В норме все компоненты комплемента - неактивные или малоактивные соединения, но могут последовательно активироваться за счет отщепления или присоединения пептидных факторов (например, С2а, C2b, C4a, C4b и др.) и факторов активации (факторов В и D, липополисахаридов, гликолипидов, антител и др.) - продукт одной реакции катализирует последующую. Катаболизм компонентов комплемента самый высокий по сравнению с другими белками сыворотки крови, с обновлением втечение суток до 50 % белков системы комплемента.
Рис. 1 . Молекула Clq -компонента комплемента (электронная микроскопия)
Молекула состоит из шести терминальных субъединиц, соединенных центральной единицей (из Schaechter M., Medoff G., Eisenstein В. Mechanisms of microbial diseases, 2 nd ed, Williams & Wilkins, 1993)
Различные компоненты комплемента и их фрагменты, образующиеся в процессе активации, способны вызывать воспалительные процессы, лизис клеток, стимулировать фагоцитоз. Конечным результатом активации может быть сборка комплекса из С5-, С6-, С7-, С8- и С9-компонентов, атакующего мембрану с образованием в ней каналов и повышением проницаемости мембраны для воды и ионов, что обусловливает гибель клетки.
Активация комплемента может происходить двумя основными путями: альтернативным - без участия антител и классическим - с участием антител (рис. 2).


Альтернативный путь более древний. В его основе лежит способность некоторых микроорганизмов активировать СЗ-конвертазу (СЗЬВЬ) путем связывания ее на углеводных участках своей поверхностной мембраны с последующей стабилизацией СЗ-кон-вертазы белком пропердином (Р). Про-пердин способен связываться с поверхностью бактериальной клетки и инициировать при этом фиксацию на ней СЗ-конвертазы и присоединение к комплементу дополнительных молекул СЗЬ. СЗЬ способен закрепляться как на поверхности микроорганизма, так и на рецепторах фагоцитов (нейтрофилов и макрофагов), выполняя роль опсонина, усиливающего фагоцитоз различных бактерий. Образовавшийся комплекс СЗЬВЬР обладает функцией СЗ-конвертазы. Формирование СЗ/С5-конвертаз при альтернативном пути активации комплемента происходит при участии факторов В, D, Р в присутствии ионов Mg 2+ и регулируется некоторыми факторами (Н, I и др.) инактивации.
Стабилизированная на мембране активная конвертаза расщепляет СЗ - один из компонентов системы комплемента, содержащийся в крови в наибольшей концентрации, что ведет к цепной реакции активации других компонентов комплемента.
В результате действия СЗ/С5-кон-вертаз вначале при участии СЗ-конвертазы происходит расщепление СЗ-компонента, содержащегося в крови в наибольшей концентрации, что ведет к цепной реакции активации других компонентов комплемента, а последующее сформирование С5-конвертазы ведет к расщеплению С5-компонента на более крупный (С5Ь) и мелкий (С5а) фрагменты. С5Ь связывается с комплексом компонентов комплемента на клеточной мембране, а С5а остается в жидкой фазе, обладая хемотаксической и анафилактогенной активностью.
С5Ь-фрагмент обладает способностью связывать компонент С6 с образованием комплекса С5Ь - С6, к которому быстро присоединяется С7, а затем С8. Комплекс С5Ь - С6, 7, 8 проникает внутрь липидного бислоя мембраны. На завершающем этапе к С8 присоединяются 12-20 молекул С9, что завершает формирование высокоактивного литического комплекса (А. А. Ярилин, 1999), формирующего трансмембранный канал, через который внутрь клетки поступают ионы водорода, натрия и вода, что ведет к набуханию и лизису клетки. С9-белок, гомологичный перфорину, способный к полимеризации при контакте с фосфолипидами мембраны, ответствен за формирование трансмембранного канала цилиндрической формы, наружная поверхность которого образована гидрофобными, а внутренняя (обращенная в полость канала) - гидрофильными участками.
Классический путь активации комплемента возник для усиления фагоцитоза в отношении микроорганизмов, которые не запускают альтернативный путь, т. е. не имеют на мембране полисахаридного участка связывания СЗ-конвертазы. Главная особенность этого пути - взаимодействие антигена и антитела с образованием иммунного комплекса (АГ-AT), активирующего компоненты комплемента (С1, С2, С4), которые, в свою очередь, формируют СЗ-конвертазу (С4Ь2а), расщепляющую СЗ-компонент.
В Сн4-доменах IgM и Сн2-доменах IgG имеются участки, обладающие сродством к Clq (только в составе иммунных комплексов). Clq связывается не менее чем с двумя Сн4-доменами одной и той же молекулы IgM и с Сн2-до-менами одновременно двух молекул IgG, в связи с чем комплементактиви-рующая активность IgG ниже, чем у IgM. С комплементсвязывающими участками антител (IgM, IgGl, IgG3 и IgG2) взаимодействуют концевые (глобулярные) участки Clq, что ведет к активации молекулы Clq, приобретающей свойства сериновой пептидгидро-лазы. Clq-пептидгидролаза активирует Clr, который участвует в активации Cls. В результате образующиеся при активации и расщеплении Clr- и Cls-фрагменты встраиваются в Clq, располагаясь между его глобулярными участками (головками). При этом формируется комплекс Clqrs, обладающий активностью трипсиновой пептидгидролазы, катализирующей расщепление С4 (на С4а- и С4Ь-фрагменты) и С2 (на С2а- и С2Ь-фрагменты). Следствием взаимодействия Clqrs, C4b и С2а в присутствии ионов Са 2+ является образование комплекса С4Ь2а, обладающего свойствами и активностью СЗ-конвертазы, расщепляющей СЗ, и участвующего в формировании С5-конверта-зы (С4Ь2аЗЬ). Дальнейшая активация комплемента по классическому пути полностью совпадает с альтернативным путем и завершается формированием мембраноатакующего комплекса С5Ь - 6789 и лизисом клеток.

Рис. 3. Аналогичные этапы активации комплемента по классическому, лектиновому и альтернативному механизмам:
Как классический так и альтернативный путь активации комплемента приводит к появлению СЗ-конвертазы: С4Ь2а и C3bBb соответственно. Классический путь начинается с активации комплексом антиген-антитело и последующего расщепления активированным CIs компонентов С4 и С2. Фрагменты меньшего размера C4a и C2b, высвобождаются, а более крупные образуют С4Ь2а. Компоненты С4 и С2 могут быть активированы также МАСП (маннан-связывающей лектин-ассоциированной сериновой протеиназой) - белком лектинового пути, аналогичным CIs, и МСЛ (сывороточным маннан-связывающим лектином). На первых этапах альтернативного пути возникший в результате «холостой» активации и связавшийся с поверхностью белок СЗЬ соединяется с фактором В, от которого Фактор D отщепляет меньший фрагмент- Ва. Больший фрагмент, то есть ВЬ, остается связанным с СЗЬ, образуя СЗЬDЬ-СЗ-конвертазу которая расщепляет дополнительное количество молекул СЗ (механизм положительной обратной связи). Поверхность активирующая комплемент (например, микроорганизмов), стабилизирует СЗЬ, обеспечивая его связывание с Фактором В Это способствует дальнейшей альтернативной активации комплемента. СЗ-конвертазы классического и альтернативного путей могут дополнительно присоединять СЗЬ, образуя ферментные комплексы, называемые С5-конвертазами (С4Ь2аЗЬ и СЗЬВЬЗЬ соответственно), которые активируют следующий компонент систем комплемента - С5 (А. Ройт и соавт., 2000)
Таким образом, принципиальных биохимических различий между классическим и альтернативным путями активации комплемента, по существу, нет, тем более что факторы В и С2, участвующие в активации СЗ по альтернативному и классическому путям, сходны между собой (по размеру, строению, фрагментам расщепления, механизму действия). Есть мнение, что, возможно, факторы В и С2 возникли в результате дубликации одного гена (В. В. Чиркин и др., 1999). Однако по клиническим проявлениям различия между этими путями весьма существенны. При альтернативном пути в циркуляторном русле значительно увеличивается содержание осколков белковых молекул с высокой биологической активностью, для нейтрализации которых включаются сложные механизмы, что повышает возможность развития вялотекущего, зачастую генерализованного воспалительного процесса. Классический путь наиболее безвреден для организма. При нем на микроорганизмы одновременно воздействуют и фагоциты, и антитела, которые специфически связывают антигенные детерминанты микроорганизмов и активизируют систему комплемента, способствуя тем самым активации фагоцитоза. При этом уничтожение атакуемой клетки происходит одновременно при участии и антител, и комплемента, и фагоцитов, что внешне может никак не проявляться. В связи с этим классический путь активации комплемента считается более физиологическим путем обезвреживания и утилизации антигенов, чем альтернативный.
Помимо двух основных путей возможны и другие механизмы активации комплемента. В частности, существует вариант классической активации комплемента - лектиновый путь активации (рис. 3), который можно трактовать и как самостоятельный (А. А. Ярилин и др., 1999; А. Ройт и соавт., 2000). Как известно, лектинами называют белки, способные специфически соединяться с определенными группами углеводов. Запуск лектиново-го пути активации комплемента связан с одним из лектинов - маннозосвязывающим белком (МСБ, содержится в сыворотке крови в концентрации 0,1 - 5,0мкг/мл). МСБ имеет очень сходную с Clq структуру хотя негомологичен ему; является Са - зависимым, обладает сродством к маннозе, которая в свободной форме присутствует на микробных клетках, но не на клетках макроорганизма. Связавшись с маннозосодержащей клеткой, МСБ приобретает способность, подобно Clqrs, активировать С4 и С2.
Дальше лектиновый и классический пути активации совпадают (А. А. Яри лин, 1999). Возможно, что лектиновый путь активации комплемента в филогенезе появился позднее альтернативного, но раньше классического. В отличие от альтернативного лектиновый путь, как и классический, включает активацию С4 и С2, но без участия антител, а с участием лишь одного МСБ. Не исключено, что появление в процессе эволюции Clq, подобного маннозосвязывающему белку, но способного приобретать активность пептидгидролазы, инициирующей каскад реакций активации комплемента лишь после взаимодействия с антигенами, привело к возникновению более эффективного классического пути активации комплемента, что значительно расширило возможности для активации комплемента у позвоночных.
Классический путь активации комплемента может также запускаться С -реактивным белком, комплексом гепарина и протамина, некоторым гликолипидами, пептидгидролазами при некоторых формах острой воспалительной реакции (пепсином, трипсином, калликреином, лизосомальными и бактериальными ферментами) на какой-либо стадии от С1 до С5.
Список литературы:
Воронин Е.С., Петров А.М., Серых М.М., Девришов Д.А. – Иммунология /Под ред. Е.С. Воронина. – М.: Колос-Пресс, 2002. – 408с.
Кульберг А.Я. /Учебное пособие – Молекулярная иммунология – М.: Высш. Шк., 1985. – 287с.
Комплемент является одним из важных факторов гуморального иммунитета, играющим роль в защите организма от антигенов. Комплемент представляет собой сложный комплекс белков сыворотки крови, находящийся обычно в неактивном состоянии и активирующийся при соединении антигена с антителом или при агрегации антигена.
1. Девять белков, составляющих собственно комплемент и обозначаемых поэтому буквой С: С1...С9, причем С1-компонент состоит из трех белковых субъединиц (С1q, С1г, С1s), все остальные представляют собой единичные белковые молекулы. В составе молекулы имеется рецептор для связывания с Рс-фрагментом молекулы антитела. Антитела, относящиеся к иммуноглобулинам различных классов, взаимодействуют с комплементами с различной степенью активности. Белки С5, С6, С7, С8 и С9 участвуют в организации мембрано-атакующего комплекса.
2. Регуляторные белки: С1Е1, С4bр, фактор Н, фактор I (инактиватор СЗb/С4b), белок S.
3. Факторы, участвующие в альтернативном пути активации системы комплемента: фактор В (протеиназа), фактор В (гликопротеин), фактор Р (пропердин) - у-глобулин, его обнаружил в 1954 г. Л. Пиллемер. Этот белок, образуя комплекс с эндотоксином, в присутствии ионов Mg разрушает С3, поэтому был назван пропердином. Пропердин стабилизирует СЗ-конвертазу альтернативного пути.
Функции комплемента многообразны: а) участвует в лизисе микробных и других клеток (цитотоксическое действие); б) обладает хемотаксической активностью; в) принимает участие в анафилаксии; г) участвует в фагоцитозе. Следовательно, комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов (например, опухолевых клеток, трансплантата).
Механизм активации комплемента представляет собой каскад ферментативных протеолитических реакций, в результате которого образуется активный цитолитический комплекс, разрушающий стенку бактерии и других клеток. Известны три пути активации комплемента: классический, альтернативный и лектиновый.
По классическому пути комплемент активируется комплексом антиген-антитело. Для этого достаточно участия в связывании антигена одной молекулы IgM или двух молекул IgG. Процесс начинается с присоединения к комплексу АГ+АТ компонента С1, который распадается на субъединицы Clq, Clr и Сls. Далее в реакции участвуют последовательно активированные «ранние» компоненты комплемента в такой последовательности: С4, С2, СЗ. Эта реакция имеет характер усиливающегося каскада, т. е. когда одна молекула предыдущего компонента активирует несколько молекул последующего. «Ранний» компонент комплемента С3 активирует компонент С5, который обладает свойством прикрепляться к мембране клетки. На компоненте С5 путем последовательного присоединения «поздних» компонентов С6, С7, С8, С9 образуется литический или мембраноатакующий комплекс который нарушает целостность мембраны (образует в ней отверстие), и клетка погибает в результате осмотического лизиса.
Альтернативный путь активации комплемента проходит без участия антител. Этот путь характерен для защиты от грамотрицательных микробов. Каскадная цепная реакция при альтернативном пути начинается с взаимодействия антигена (например, полисахарида) с протеинами В, D и пропердином (Р) с последующей активацией компонента СЗ. Далее реакция идет так же, как и при классическом пути - образуется мембраноатакующий комплекс.
Лектиновыи путь активации комплемента также происходит без участия антител. Он инициируется особым маннозосвязывающим белком сыворотки крови, который после взаимодействия с остатками маннозы на поверхности микробных клеток катализирует С4. Дальнейший каскад реакций сходен с классическим путем.
В процессе активации комплемента образуются продукты протеолиза его компонентов - субъединицы СЗа и СЗb, С5а и С5b и другие, которые обладают высокой биологической активностью. Например, СЗа и С5а принимают участие в анафилактических реакциях, являются хемоаттрактантами, СЗb - играет роль в опсонизации объектов фагоцитоза, и т. д. Сложная каскадная реакция комплемента происходит с участием ионов Са 2+ и Mg 2+ .
1) способность связываться с комплексом антиген + антитело;
2) лизирование эритроцитов, использованных для получения гемолитической сыворотки.
РСК ставят в два этапа, и в ней соответственно участвуют две системы - опытная, или диагностическая, и индикаторная. Диагностическая система состоит из исследуемой (или диагностической) сыворотки, которую перед постановкой реакции прогревают при 56 °С в течение 30 мин для инактивации имеющегося в ней комплемента, и антигена. К этой системе добавляют стандартный комплемент. Его источником служит свежая или высушенная сыворотка морской свинки. Смесь инкубируют при 37 "С в течение одного часа. Если в исследуемой сыворотке имеются антитела, произойдет их взаимодействие с добавленным антигеном, и образующиеся комплексы антиген + антитело свяжут добавленный комплемент. Если же в сыворотке антитела отсутствуют, образования комплекса антиген + антитело не произойдет, и комплемент останется свободным. Никаких видимых проявлений связывания комплемента на этой стадии реакции обычно нет. Поэтому для выяснения вопроса, произошло или нет связывание комплемента, добавляют вторую, индикаторную систему (инактивированная гемолитическая сыворотка + эритроциты барана), и смесь всех компонентов РСК вновь инкубируют при 37С в течение 30-60 мин, после чего оценивают результаты реакции. В случае, если комплемент связался на первой стадии, в диагностической системе, т. е. в сыворотке больного имеются антитела, и произошло связывание комплемента комплексом антитело + + антиген, лизиса эритроцитов не будет - РСК положительна: жидкость бесцветна, на дне пробирки осадок эритроцитов. Если же в сыворотке специфические антитела отсутствуют и связывания комплемента в диагностической системе не произойдет, т. е. РСК отрицательна, то неизрасходованный в диагностической системе Комплемент связывается с комплексом эритроциты + антитела индикаторной системы и произойдет гемолиз: в пробирке «лаковая кровь», осадка эритроцитов нет. Интенсивность РСК оценивают по четырехкрестной системе в зависимости от степени задержки гемолиза и наличия осадка эритроцитов. Реакция сопровождается соответствующими контролями: контроль сыворотки (без антигена) и контроль антигена (без сыворотки), так как некоторые сыворотки и некоторые антигены обладают антикомплементарным действием. Перед постановкой РСК все компоненты, участвующие в ней, за исключением исследуемой сыворотки или антигена, подвергаются тщательному титрованию. Особенно важно ввести в реакцию точную дозу комплемента, так как его нехватка или избыток могут привести к ложным результатам. Титром комплемента является то его минимальное количество, которое в присутствии рабочей дозы гемолитической сыворотки обеспечивает полное растворение эритроцитов. Для постановки основного опыта берут дозу комплемента, увеличенную на 20-25 % по сравнению с установленным титром. Титром гемолитической сыворотки является то ее максимальное разведение, которое, будучи смешано с равным объемом 10 % раствора комплемента, полностью гемолизирует соответствующую дозу эритроцитов в течение 1 ч при температуре 37 °С. В основной опыт берут сыворотку, разведенную до 1/3 своего титра.
Приобретенный иммунитет. Значение антител в формировании иммунитета. Роль различных классов иммуноглобулинов в иммунологических реакциях (агглютинации, РСК, нейтрализации токсинов и вирусов, развитии местного иммунитет).
Приобретенный иммунитет отличается от видового следующими особенностями.
Во-первых, он не передается по наследству. По наследству передается лишь информация об органе иммунитета, а сам иммунитет формируется в процессе индивидуальной жизни в результате взаимодействия с соответствующими возбудителями или их антигенами.
Во-вторых, приобретенный иммунитет является строго специфическим, т. е. всегда направлен против конкретного возбудителя или антигена.
Формирование приобретенного специфического иммунитета происходит благодаря кооперативному взаимодействию макрофагов (и других антигенпредставляющих клеток), В- и Т-лимфоцитов и при активном участии всех остальных иммунных систем.
Одним из характерных признаков приобретенного иммунитета служит появление в сыворотке крови и тканевых соках специфических защитных веществ - антител, направленных против чужеродных веществ.
Антитела являются уникальными сывороточными белками - глобулинами, которые вырабатываются в ответ на поступление в организм антигена и способны с ним специфически взаимодействовать. Совокупность сывороточных белков, обладающих свойствами антител, называют иммуноглобулинами и обозначают символом Ig.
Существует пять различных классов иммуноглобулинов: IgG, IgМ, IgА, IgЕ, IgD Они различаются по молекулярной массе, содержанию углеводов, составу полипептидных цепей, коэффициентам седиментации и др.
Высокая нейтрализующая активность антител, принадлежащих к IgG, свидетельствует о важной роли их в антитоксическом иммунитете. Антитела IgМ особенно активны в реакциях фагоцитоза с корпускулярными антигенами и поэтому играют существенную роль в антимикробном иммунитете, В реакциях нейтрализации вирусов особенно активны антитела IgА, следовательно, им принадлежит большая роль в противовирусном иммунитете. Кроме того, секреторные IgAs обусловливают местный иммунитет слизистых оболочек. Наконец, антитела IgЕ, обладающие гомоцитотропностью, опосредуют реакции гиперчувствительности немедленного типа.
Реакция связывания комплемента.
Уникальная способность комплемента специфически связываться с различными по своей природе комплексами антиген + антитело нашла широкое применение в реакции связывания комплемента (РСК). Особое преимущество РСК состоит в том, что природа антигена, участвующего в ней (корпускулярный или растворимый), не имеет значения, так как комплемент связывается с Fс-фрагментом любого антитела, относящегося к IgG и IgМ, независимо от его антительной специфичности. Кроме того, РСК очень чувствительна: она позволяет обнаружить количество антител в 10 раз меньшее, чем, например, в реакции преципитации. РСК была предложена в 1901 г. Ж. Борде и О. Жангу. В ее основе лежат два свойства комплемента:
3) способность связываться с комплексом антиген + антитело;
4) лизирование эритроцитов, использованных для получения гемолитической сыворотки.
РСК ставят в два этапа, и в ней соответственно участвуют две системы - опытная, или диагностическая, и индикаторная. Диагностическая система состоит из исследуемой (или диагностической) сыворотки, которую перед постановкой реакции прогревают при 56 °С в течение 30 мин для инактивации имеющегося в ней комплемента, и антигена. К этой системе добавляют стандартный комплемент. Его источником служит свежая или высушенная сыворотка морской свинки. Смесь инкубируют при 37С в течение одного часа. Если в исследуемой сыворотке имеются антитела, произойдет их взаимодействие с добавленным антигеном, и образующиеся комплексы антиген + антитело свяжут добавленный комплемент. Если же в сыворотке антитела отсутствуют, образования комплекса антиген + антитело не произойдет, и комплемент останется свободным. Никаких видимых проявлений связывания комплемента на этой стадии реакции обычно нет. Поэтому для выяснения вопроса, произошло или нет связывание комплемента, добавляют вторую, индикаторную систему (инактивированная гемолитическая сыворотка + эритроциты барана), и смесь всех компонентов РСК вновь инкубируют при 37С в течение 30-60 мин, после чего оценивают результаты реакции. В случае, если комплемент связался на первой стадии, в диагностической системе, т. е. в сыворотке больного имеются антитела, и произошло связывание комплемента комплексом антитело + + антиген, лизиса эритроцитов не будет - РСК положительна: жидкость бесцветна, на дне пробирки осадок эритроцитов. Если же в сыворотке специфические антитела отсутствуют и связывания комплемента в диагностической системе не произойдет, т. е. РСК отрицательна, то неизрасходованный в диагностической системе Комплемент связывается с комплексом эритроциты + антитела индикаторной системы и произойдет гемолиз: в пробирке «лаковая кровь», осадка эритроцитов нет. Интенсивность РСК оценивают по четырехкрестной системе в зависимости от степени задержки гемолиза и наличия осадка эритроцитов. Реакция сопровождается соответствующими контролями: контроль сыворотки (без антигена) и контроль антигена (без сыворотки), так как некоторые сыворотки и некоторые антигены обладают антикомплементарным действием. Перед постановкой РСК все компоненты, участвующие в ней, за исключением исследуемой сыворотки или антигена, подвергаются тщательному титрованию. Особенно важно ввести в реакцию точную дозу комплемента, так как его нехватка или избыток могут привести к ложным результатам. Титром комплемента является то его минимальное количество, которое в присутствии рабочей дозы гемолитической сыворотки обеспечивает полное растворение эритроцитов. Для постановки основного опыта берут дозу комплемента, увеличенную на 20-25 % по сравнению с установленным титром. Титром гемолитической сыворотки является то ее максимальное разведение, которое, будучи смешано с равным объемом 10 % раствора комплемента, полностью гемолизирует соответствующую дозу эритроцитов в течение 1 ч при температуре 37 °С. В основной опыт берут сыворотку, разведенную до 1/3 своего титра.
Непрямая реакция гемолиза используется как ускоренный метод обнаружения специфических антител. В качестве носителя антигенов используют эритроциты. При наличии в сыворотке больного специфических антител сенсибилизированные эритроциты в присутствии комплемента лизируются.
Антигены. Определение понятия, свойства, химическая природа. Специфичное антигенов. Детерминантная группа (эпитоп), шлеппер. Полноценные и неполноценные антигены. Гаптены и полугаптены. Факторы, определяющие антигенность белка и ее специфичность.
Антигены - любые вещества, содержащиеся в микроорганизмах и других клетках или выделяемые ими, которые несут признаки генетически чужеродной информации и при введении в организм вызывают развитие специфических иммунных реакций.
Реализация антигенности зависит от способности антигена метаболизироваться в организме, т. е. быть объектом разрушающего действия макрофагов и взаимодействовать с другими клетками иммунной системы. Благодаря такому взаимодействию происходит распознавание антигенной специфичности. Все антигены обладают специфичностью, т. е. определенными особенностями, генетически детерминированными и связанными с их структурой, почему они и отличаются друг от друга.
Для характеристики микроорганизмов помимо родовой, видовой и групповой антигенной специфичности очень важное значение имеет определение типоспецифичности антигенов. Типоспецифичность - особенность антигенного строения, которая обусловливает различия среди особей одной группы сходных организмов данного вида и позволяет выделить среди них серотипы, или сероварианты (серовары). Выявление сероваров дает возможность осуществлять очень тонкую дифференциацию внутри вида микроорганизмов.
Изучение антигенных свойств различных сложных химических соединений - белков, полисахаридов, липидов, нуклеиновых кислот и т. д. - показало, что существует два типа антигенов - полноценные и неполноценные.
Полноценные антигены обладают обеими функциями антигена: способностью индуцировать образование антител и специфически с ними взаимодействовать.
Неполноценные антигены сами по себе способностью индуцировать образование антител не обладают, они приобретают это свойство только после соединения с белками или другими полноценными антигенами. Такие неполноценные антигены называются гаптенами или полугаптенами.
Неполноценные антигены обладают только одним свойством антигена: они способны специфически взаимодействовать с теми антителами, в индукции синтеза которых они участвовали (после присоединения к белку и превращения в полноценные антигены).
Если взаимодействие неполноценного антигена с антителом сопровождается обычными иммунологическими реакциями, его называют гаптеном. Если неполноценный антиген имеет очень небольшую молекулярную массу и его взаимодействие с антителами не сопровождается обычными видимыми реакциями, его называют полугаптеном. О присутствии полугаптена в этом случае судят по тому признаку, что антитела, будучи связаны с полугаптеном, уже не проявляют себя в обычной реакции с полноценным антигеном (задерживающая реакция Ландштейнера).
Антигенное строение микробной клетки. Основные группы антигенов. Химическая природа антигенной специфичности. Значение изучения антигенов в серологической классификации микроорганизмов.
Для медицинской микробиологии наибольший интерес представляют антигенные свойства бактерий, токсинов и вирусов. Результаты их изучения используются в практике получения высокоэффективных иммуногенных препаратов, а также для совершенствования методов идентификации возбудителей болезней.
Антигенное строение микробной клетки. Н-, О- и К-антигены, токсины и ферменты бактерий как антигены. Перекрестнореагирующие антигены. Принципы определения антигенного состава бактерий, дифференциация общих (групповых) типоспецифических антигенов.
Антигенное строение микробной клетки. Обладая сложным химическим строением, бактериальная клетка представляет собой целый комплекс антигенов. Антигенными свойствами обладают жгутики, капсула, клеточная стенка, цитоплазматическая мембрана, рибосомы и другие компоненты цитоплазмы, а также различные продукты белковой природы, выделяемые бактериями во внешнюю среду, в том числе токсины и ферменты. В связи с этим различают следующие основные виды микробных антигенов: соматические, или О - антигены; жгутиковые, или Н-антигены; поверхностные, или капсульные К-антигены.
Видовая специфичность - антигенные особенности, присущие представителям данного вида. Отпечаток видовой специфичности имеют многие макромолекулы данного организма. Определение видовых антигенов может быть использовано для дифференциации особей одного вида от другого.
Групповая специфичность - особенности антигенного строения, свойственные определенной группе особей внутри данного вида организмов. Групповые антигены, позволяющие различать отдельных особей или группы особей внутри одного вида, называются изоантигенами.
Гетероспецифичность - антигенная специфичность, обусловленная наличием общих для представителей разных видов антигенов. Гетероантигены обусловливают перекрестные иммунологические реакции.
Типоспецифичность - особенность антигенного строения, которая обусловливает различия среди особей одной группы сходных организмов данного вида и позволяет выделить среди них серотипы, или сероварианты (серова- ры). Выявление сероваров дает возможность осуществлять очень тонкую дифференциацию внутри вида микроорганизмов.
Большинство современных классификаций патогенных микроорганизмов построены с учетом этих типов антигенной специфичности.
№ 6 Комплемент, его структура, функции, пути активации, роль в иммунитете.
Природа и характеристика комплемента
.
Комплемент является одним из важных факторов гуморального иммунитета, играющим роль в защите организма от антигенов. Комплемент представляет собой сложный комплекс белков сыворотки крови, находящийся обычно в неактивном состоянии и активирующийся при соединении антигена с антителом или при агрегации антигена. В состав комплемента входят 20 взаимодействующих между собой белков, девять из которых являются основными компонентами комплемента; их обозначают цифрами: С1
, С2, СЗ, С4...С9. Важную роль играют также факторы В, D
и Р
(пропердин). Белки комплемента относятся к глобулинам и отличаются между собой по ряду физико-химических свойств. В частности, они существенно различаются по молекулярной массе, а также имеют сложный субъединичный состав: C
1-
C
1
q
,
C
1
r
,
C
1
s
; СЗ-СЗа, СЗ
b
; С5-С5а, С5
b
и т. д. Компоненты комплемента синтезируются в большом количестве (составляют 5-10% от всех белков крови), часть из них образуют фагоциты.
Функции комплемента
многообразны: а) участвует в лизисе микробных и других клеток (цитотоксическое действие); б) обладает хемотаксической активностью; в) принимает участие в анафилаксии; г) участвует в фагоцитозе. Следовательно, комплемент является компонентом многих иммунологических реакций, направленных на освобождение организма от микробов и других чужеродных клеток и антигенов
(например, опухолевых клеток, трансплантата).
Механизм активации
комплемента
очень сложен и представляет собой каскад ферментативных протеолитических реакций, в результате которого образуется активный цитолитический комплекс, разрушающий стенку бактерии и других клеток. Известны три пути активации комплемента: классический, альтернативный и лектиновый.
По классическому пути
комплемент активируется комплексом антиген-антитело. Для этого достаточно участия в связывании антигена одной молекулы
IgM
или двух молекул
IgG
. Процесс начинается с присоединения к комплексу АГ+АТ компонента С1
, который распадается на субъединицы C
1
q
,
C
1
r
и С1
s
. Далее в реакции участвуют последовательно активированные «ранние» компоненты комплемента в такой последовательности: С4
, С2, СЗ. Эта реакция имеет характер усиливающегося каскада, т. е. когда одна молекула предыдущего компонента активирует несколько молекул последующего. «Ранний» компонент комплемента С3 активирует компонент С5, который обладает свойством прикрепляться к мембране клетки. На компоненте С5 путем последовательного присоединения «поздних» компонентов С6, С7, С8, С9 образуется литический или мембраноатакующий комплекс который нарушает целостность мембраны (образует в ней отверстие), и клетка погибает в результате осмотического лизиса.
Альтернативный путь
активации комплемента проходит без участия антител. Этот путь характерен для защиты от грамотрицательных микробов. Каскадная цепная реакция при альтернативном пути начинается с взаимодействия антигена (например, полисахарида) с протеинами В
, D
и пропердином (Р) с последующей активацией компонента СЗ. Далее реакция идет так же, как и при классическом пути - образуется мембраноатакующий комплекс.
Лектиновый путь
активации комплемента также происходит без участия антител. Он инициируется особым маннозосвязывающим белком
сыворотки крови, который после взаимодействия с остатками маннозы на поверхности микробных клеток катализирует С4. Дальнейший каскад реакций сходен с классическим путем.
В процессе активации комплемента образуются продукты протеолиза его компонентов - субъединицы СЗа и СЗ
b
, С5а и С5
b
и другие, которые обладают высокой биологической активностью. Например, СЗа и С5а принимают участие в анафилактических реакциях, являются хемоаттрактантами, СЗ
b
- играет роль в опсонизации объектов фагоцитоза, и т. д. Сложная каскадная реакция комплемента происходит с участием ионов Са 2
+ и Mg
2+
.